Peptides for diabetes research
The number of cases and the prevalence of diabetes have steadily increased over the past decades. Globally, there are approximately 422 million diabetics, most of whom live in low- and middle-income countries, and every year 1.5 million deaths are directly attributed to diabetes. Diabetes mellitus can be divided into two main types, type 1 or insulin-dependent diabetes mellitus (IDDM) and type 2, or non insulin-dependent diabetes mellitus (NIDDM). The absolute lack of insulin, due to destruction of the insulin producing pancreatic β-cells, is the particular disorder in type 1 diabetes. Type 2 diabetes is mainly characterized by the inability of cells to respond to insulin. The condition affects mostly the cells of muscle and fat tissue, and results in a condition known as “insulin resistance”.
Effects of diabetes
Over time, diabetes mellitus can lead to blindness, kidney failure, and nerve damage. Diabetes mellitus is also an important factor in accelerating the hardening and narrowing of the arteries (atherosclerosis), leading to stroke, coronary heart diseases, and other blood vessel disorders.
Introduction
Diabetes was already known in ancient times. The name of this disease was created by the Graeco-Roman physician Aretaeus of Cappadocia (approx. 80 – 130 AD) and is derived from the Greek word diabainein that means ‘to flow through’. The adjective mellitus, which comes from Latin and means ‘honey-sweet’, was added by the German physician Johann Peter Frank (1745-1821).
It was introduced in order to distinguish diabetes mellitus, also called ‘sugar diabetes’, from diabetes insipidus, where an excessive amount of urine is produced as a result of a disturbance of the hormonal control of reabsorption of water in the kidneys. In 1889, pancreatic secretions were shown to control blood sugar levels. However, it took another 30 years until insulin was purified from the islets of Langerhans. In the following 50 years scientists detected the system-wide effects of insulin in liver, muscle, and adipose tissues. In the 1970s, the insulin receptor was discovered, and 10 years later, its tyrosine kinase activity was
demonstrated.
Despite this steady progress, one of the most challenging health problems of the 21st century remains the dramatic increase in diabetes mellitus that is occurring throughout the world. Today diabetes mellitus is one of the main causes of death in most developed countries. According to data from the International Diabetes Federation, more than 382 million people around the world suffered from diabetes in 2013. This alarming number could reach 592 million by 2035. Further 316 million people have impaired glucose tolerance, a condition that can signal oncoming diabetes. 85 – 95 % of the diabetics have type 2 diabetes, a chronic disease associated with insulin deficiency and insulin resistance. Complications seen with diabetes range from heart disease (2 to 4 times higher occurence than in non-diabetics) to blindness, kidney disease, amputations, nerve damage and erectile dysfunction.
As obesity spreads, the number of type 2 diabetics rises. Over 80% of diabetics are obese. Consequently, the treatment of risk factors such as obesity, hypertension, and hyperlipidemia assumes major importance and must be coordinated with a good glycemic control for the reduction in total mortality in type 2 diabetes mellitus. In this monograph, we describe the pancreatic and gastrointestinal peptide hormones that are involved in the control of blood glucose, the classification, and the treatment of diabetes mellitus.
Pancreatic Peptide Hormones
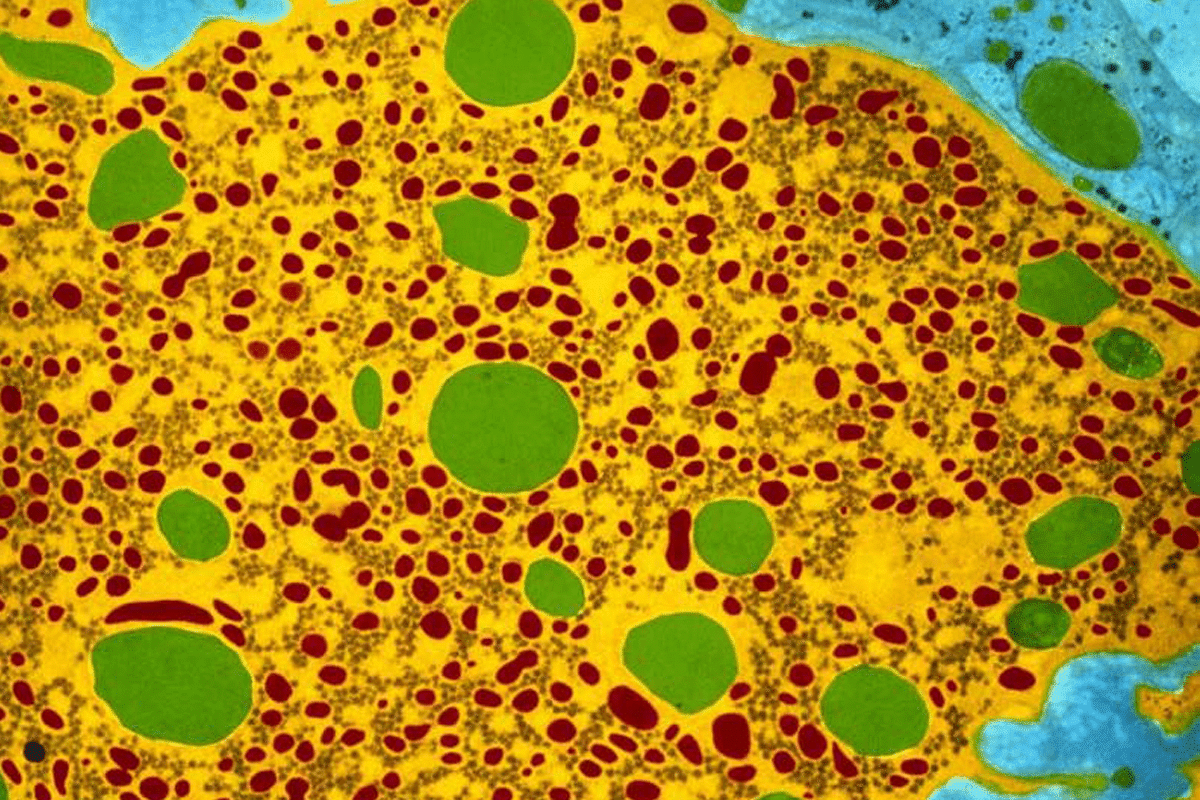
The islets of Langerhans contain four main cell types: β-cells secreting insulin, α-cells secreting glucagon, δ-cells secreting somatostatin and γ-cells secreting pancreatic polypeptide (PP). The core of each islet contains mainly the β-cells surrounded by a mantle of α-cells interspersed with δ-cells or γ-cells. Insulin is synthesized as a preprohormone in the β-cells of the islets of Langerhans. Removal of its signal peptide during insertion into the endoplasmic reticulum generates proinsulin which consists of 3 domains: an amino-terminal B-chain, a carboxy-terminal A-chain and a connecting peptide known as C-peptide. Within the endoplasmic reticulum proinsulin is exposed to several specific endopeptidases.
These enzymes excise the C-peptide, thereby generating the mature form of insulin, a small protein consisting of an A-chain of 21 amino acids (containing an internal disulfide bridge) linked by two disulfide bridges to a B-chain of 30 amino acids. β-Cells secrete insulin in response to a rising levelbof circulating glucose. The normal fasting blood glucose concentration in humans and most mammals is 80 to 90 mg per 100 ml, associated with very low levels of insulin secretion. After a meal, excess sugars must be stored so that energy reserves will be available later on. Excess glucose is sensed by β-cells in the pancreas, which respond by secreting insulin into the bloodstream. Insulin causes various cells in the body to store glucose (see Fig. 1):
- Insulin stimulates skeletal muscle fibers to convert glucose into glycogen. It also induces the synthesis of proteins from amino acids circulating in the blood.
- Insulin acts on liver cells. It stimulates them to take up glucose from the blood converting it into glycogen while inhibiting the production of the enzymes involved in glycogenolysis.
- Insulin acts on fat cells to stimulate the uptake of glucose and the synthesis of fat. In each case, insulin triggers these effects by binding to the insulin receptor, a heterotetramer of two extracellular α-subunits that are bonded by disulfides to two transmembrane β-subunits. Insulin receptor activation leads to specific phosphorylation events followed by an increase in glucose storage and a concomitant decrease in hepatic glucose release.
C-Peptide is applied as a diagnostic tool. It is released in amounts equal to insulin, so the level of C-peptide in the blood indicates how much insulin is being produced by the pancreas. The concentration of C-peptide is measured in diabetics to differentiate between endogenous (produced by the body) and exogenous (injected into the body) insulin, since synthetic insulin does not contain the C-peptide. Inappropriate use of insulin in persons with a low blood sugar level results in a low C-peptide level.
Glucose homeostasis is accomplished by complex physiological mechanisms. Control of blood glucose levels involves insulin, glucagon and other peptide hormones such as glucagon-like peptide 1 (GLP-1) and glucosedependent insulinotropic polypeptide (gastric inhibitory polypeptide (GIP)).
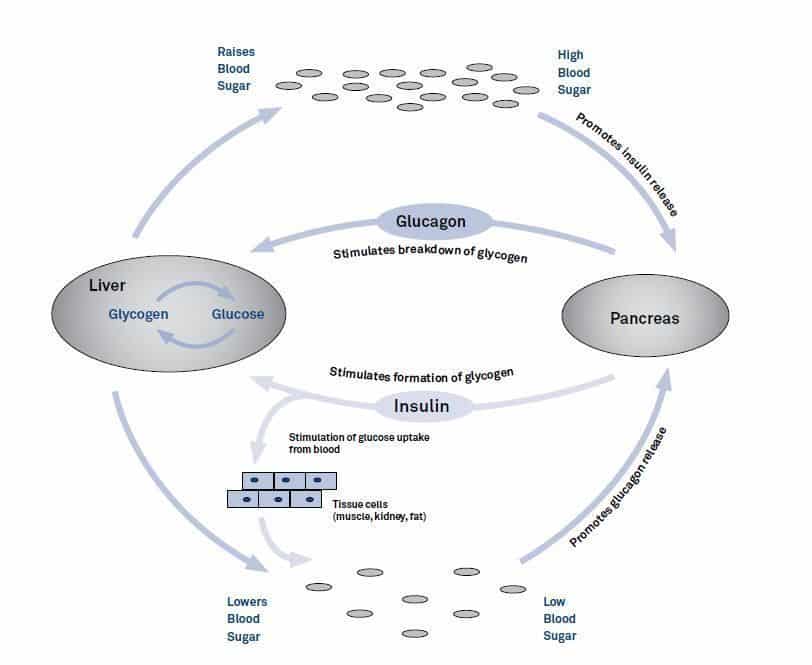
Fig. 1. Opposing effects of insulin and glucagon
The C-peptide level can also be determined in patients with type 2 diabetes showing how much insulin is produced by the β-cells. Abnormal high amounts of C-peptide can indicate the presence of a tumor called insulinoma which secretes insulin. β-Cells also secrete a peptide hormone known as islet amyloid polypeptide (IAPP) or amylin.
This 37 amino acid peptide is structurally related to calcitonin and has weak calcitonin-like effects on calcium metabolism and osteoclast activity. Amylin shows about 50% sequence identity with calcitonin gene-related peptide (CGRP). It is stored together with insulin in the secretory granules of β-cells and is co-secreted with insulin. Amylin’s most potent actions include the slowing of gastric emptying and the suppression of postprandial glucagon secretion. The hormone also reduces food intake and inhibits the secretion of gastric acid and digestive enzymes. Thus, there is therapeutic potential of IAPP agonists for the treatment of patients with absolute amylin deficiency (type 1 diabetes) or relative amylin deficiency (type 2 diabetes).
In addition, amylin is the major component of the pancreatic amyloid deposits occurring in the pancreas of patients with type 2 diabetes. Glucagon secretion is stimulated by low, and inhibited by high concentrations of glucose and fatty acids in the plasma (see Fig. 1). It counterbalances the action of insulin, increasing the levels of blood glucose and stimulating the protein breakdown in muscle.
Glucagon is a major catabolic hormone, acting primarily on the liver. The peptide stimulates glycogenolysis (glycogen breakdown) and gluconeogenesis (synthesis of glucose from non carbohydrate sources), inhibits glycogenesis (glycogen synthesis) and glycolysis, overall increasing hepatic glucose output and ketone body formation. In people suffering from diabetes, excess secretion of glucagon plays a primary role in hyperglycemia (high blood glucose concentration). Glucagon is clinically used in the treatment of hypoglycemia in unconscious patients (who can’t drink).
Somatostatin release from the pancreas and gut is stimulated by glucose and amino acids. In diabetes, somatostatin levels are increased in pancreas and gut, presumably as a consequence of insulin deficiency. Somatostatin inhibits secretion of growth hormone, insulin and glucagon.
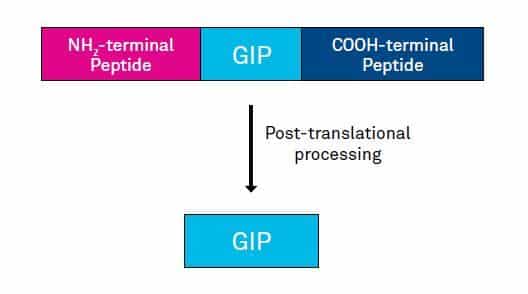
Fig. 2. Structure of prepro-GIP
Gastrointestinal Peptide Hormones
Glucose-dependent insulinotropic polypeptide (GIP) and glucagon-like peptide 1 (GLP-1) have significant effects on insulin secretion and glucose regulation. They are released after ingestion of carbohydrateand fat-rich meals and stimulate insulin secretion postprandially. Both gut hormones constitute the class of incretins and share considerable sequence homology.
GIP is a single 42 amino acid peptide derived from a larger 153 amino acid precursor (see Fig. 2). The peptide was originally observed to inhibit gastric acid secretion (hence it was designated gastric inhibitory polypeptide). Subsequent studies have demonstrated potent glucose-dependent insulin stimulatory effects of GIP administration in dogs and rodents.
GIP also regulates fat metabolism in adipocytes, including stimulation of lipoprotein lipase activity, fatty acid incorporation, and fatty acid synthesis. Unlike GLP-1, GIP does not inhibit glucagon secretion or gastric emptying. The peptide promotes β-cell proliferation and cell survival in islet cell line studies. GLP-1 is derived from the product of the proglucagon gene. This gene encodes a preproprotein (see Fig. 3) that is differentially processed dependent on the tissue in which it is expressed. In pancreatic α-cells, prohormone convertase 2 action leads to the release of glucagon.
In the gut, prohormone convertase 1/3 action leads to the release of several peptides including GLP- 1. Bioactive GLP-1 consists of two forms: GLP-1 (7-37) and GLP-1 (7-36) amide. The latter form constitutes the majority (80%) of the circulating GLPs.
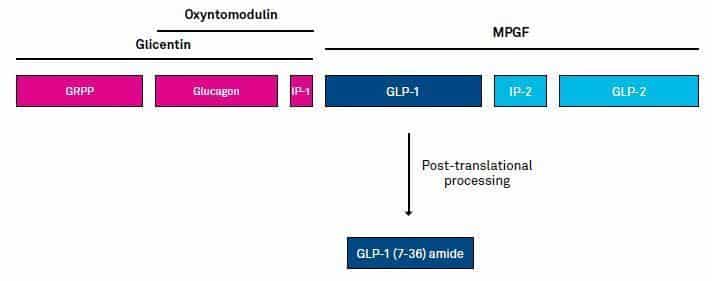
Fig. 3. Structure of preproglucagon: GRPP, glicentinrelated pancreatic peptide; IP, intervening peptides. Further peptides derived from the preproprotein include glicentin which is composed of amino acids 1-69, oxyntomodulin (glucagon-37) consisting of amino acids 33-69, and the major proglucagon fragment (MPGF) comprising amino acids 72-158.
The primary physiological responses to GLP-1 are glucose-dependent insulin secretion, inhibition of glucagon secretion and inhibition of gastric acid secretion and gastric emptying. All effects of GLP-1 are exerted by activation of the GLP-1 receptor, a seven transmembrane spanning Gprotein-coupled receptor (GPCR), leading to increased cAMP production and enhanced protein kinase A (PKA) activity.
The potential use of GLP-1 for the treatment of diabetes has been considered. GLP-1 exerts antidiabetogenic properties in subjects with type 2 diabetes by stimulating insulin secretion, increasing β-cell mass, inhibiting glucagon secretion, delaying gastric emptying, and inducing satiety, thus slowing the entry of sugar into the blood. However, GLP-1 is rapidly degraded by the enzyme dipeptidyl peptidase IV (DPP IV), making it unattractive as a therapeutic agent. Successful strategies to overcome this difficulty are the use of DPP IV-resistant GLP-1 receptor agonists, such as liraglutide (NN2211, a fatty acid-linked DPP IV-resistant derivative of GLP-1) or exendin-4 (ex-enatide).
An alternative approach is the use of inhibitors of DPP IV, such as sitagliptin, P32/98 (H-Ile-thiazolidide hemifumarate), NVP DPP 728 (N-[2-(5-Cyanopyridin-2-ylamino) ethyl]-Gly-Pro-nitrile) and the Gly- Pro-nitrile-derived compounds vildagliptin and saxagliptin. Exendin-4 is a peptide hormone found in the saliva of the Gila monster, a lizard native to several Southwestern American states. Like GLP-1 exendin-4 exerts its effects through the GLP-1 receptor but is much more potent than GLP-1. Exenatide, a synthetic form of exendin-4 has been approved by the FDA as an antidiabetic drug.
In contrast to most drugs that work by only one mechanism, exendin-4 acts by multiple mechanisms, such as stimulation of insulin secretion, slowing gastric emptying, and inhibiting the production of glucose by the liver. Furthermore, exendin-4 was shown to suppress appetite and promote weight loss.
Classification of Diabetes Mellitus
The International Diabetes Federation distinguishes between three main types of diabetes mellitus. This division is based upon whether the ‘blood sugar problem‘ is caused by insulin deficiency or insulin resistance: Type 1 diabetes (formerly known as insulindependent diabetes mellitus or juvenileonset diabetes) is a β-islet cell specific, T-lymphocyte-mediated autoimmune disorder. It is characterized by a failure of the pancreas to produce sufficient insulin.
Without insulin to promote the cellular uptake of glucose, the blood glucose concentrations reach high levels. At concentrations above 10 mM, renal tubular reabsorption is saturated and glucose is passed into the urine.
The classic symptoms are excessive secretion of urine, thirst, weight loss and tiredness. It is known that multiple genes contribute to the familial clustering of this disease, the major histocompatibility complex (MHC) being the most important of these. The MHC class 2 genotype is one of the strongest genetic factors determining disease susceptibility.
Type 2 diabetes (formerly named noninsulin-dependent diabetes mellitus ormaturity-onset diabetes) is associated with insulin resistance rather than the lack of insulin as seen in type 1 diabetes. This lack of insulin sensitivity results in higher than normal blood glucose levels. Type 2 diabetes is not HLA-linked and no autoimmune destruction of the pancreatic cells is observed. The development of type 2 diabetes seems to be multifactorial.
Genetic predisposition appears to be the strongest factor. Other risk factors are obesity and high caloric intake. Pancreatic α-cell mass is increased, followed by an exaggerated response of glucagon to amino acids and an impaired suppression of glucagon secretion by hyperglycemia. Increased hepatic production of glucose with a failure of the pancreas to adapt to this situation and resistance to the action of insulin are characteristic features of this disorder. Another important morphological feature is the amyloid deposition in islets.
These deposits consist of islet amyloidpolypeptide or amylin, that is believed, to originate in the β-cell secretory granule. Type 2 diabetes occurs most frequently in adults, but is being noted increasingly in adolescents as well. Type 2 diabetes develops slowly and the symptoms are usually less severe than in type 1. Sometimes the disease is only diagnosed several years after its onset, when complications are already present.
Common late microvascular complications include retinopathy, nephropathy, and peripheral and autonomic neuropathies. Macrovascular complications include atherosclerotic coronary peripheral
arterial disease. Gestational diabetes may be observed in non-diabetic women during late pregnancy. They develop a resistance to insulin and, subsequently, high blood glucose.
This type of diabetes is probably caused by hormones produced by the placenta. The blood glucose level has to be carefully controlled as to minimize risks for mother and child, it will return to the normal value after birth.
Advances in genomics, proteomics and metabolomics will help us to further understand the causes of type 1 and type 2 diabetes and might eventually lead to novel therapeutic approaches.
Treatment of Diabetes Mellitus
Insulin is essential for the treatment of type 1 diabetes. The effects of insulin and its mechanism of action are described above. For clinical application, either porcine or bovine insulin was given formerly. Today, human insulin (produced recombinantly) is used. A new approach is the production of orally active insulin using modifications to make insulin resistant to enzymatic breakdown, facilitating absorption.
Most of the vascular consequences of insulin resistance are due to the hyperglycemia seen in type 2 diabetes. For this reason a major goal of therapeutic intervention in type 2 diabetes is to reduce circulating glucose levels. There are many pharmacological strategies to accomplish these goals:
- The use of α-glucosidase inhibitors (e.g. acarbose) leads to a reduction in digestion and thereby minimizes the consequent absorption of glucose into the systemic circulation. The reduction in glucose uptake allows the pancreatic β-cells to regulate the insulin secretion more effectively. The advantage of α-glucosidase inhibitors is that they function locally in the intestine and have no major systemic action. Plants are rich sources of α-glucosidase inhibitors, some of which are being evaluated for their therapeutic potential.
- The sulfonylureas (e.g. glibenclamide) are referred to as endogenous insulin secretagogues because they induce the pancreatic release of insulin and thus reduce plasma glucose. Sulfonylureas function by binding to and inhibiting the pancreatic ATP-dependent potassium channels normally involved in the glucose-mediated insulin secretion. Unwanted side-effects of sulfonylureas are appetite stimulation, probably via their effects on insulin secretion and blood glucose, often leading to weight gain.
- The biguanides (e.g. metformin) are a class of drugs that lower blood glucose levels by enhancing insulin-mediated suppression of hepatic glucose production (gluconeogenesis) and by enhancing insulin-stimulated glucose uptake by skeletal muscle. Metformin is currently the most widely prescribed insulin-sensitizing drug in clinical use. The major site of action for metformin is the liver. Its use can be contraindicated in patients with liver dysfunction.
- The thiazolidinediones (e.g. pioglitazone) have been proven useful in treating the hyperglycemia associated with insulin resistance in both type 2 diabetes and non-diabetic conditions. These products function as agonists for the peroxisome proliferator-activated receptor-γ (PPAR-γ). PPARs are members of a nuclear receptor superfamily that has important roles in carbohydrate and lipid metabolism. Thiazolidinediones enhance peripheral sensitivity to insulin and, to a lesser degree, decrease hepatic glucose production by binding to and activating the PPAR-γ. Adverse effects of thiazolidine-diones include weight gain, anemia, and abnormalities in liver and enzyme levels. Resistin, an adipocyte-derived peptide, first identified during a search for targets of thiazolidinediones, has been found to be downregulated by thiazolidinediones.
- GLP-1 analogs stimulate insulin release, inhibit glucagon secretion, slow gastric emptying and stimulate β-cell proliferation. One of the most promising GLP-1 receptor agonists is exenatide (exendin-4) which is 53% identical to human GLP-1 at the amino acid level. The main advantage of exenatide is its resistance to cleavage and inactivation by dipeptidyl-peptidase IV (DPP IV). In 2005, the FDA approved exenatide as adjunctive therapy to improve blood sugar control in patients with type 2 diabetes who have not achieved adequate control with metformin and/or a sulfonylurea. Use of the peptide as primary monotherapy was approved in 2009, and a sustained release formulation in 2014. The long-acting GLP-1 agonist liraglutide was approved by the authority for the treatment of type 2 diabetes in 2010. Albiglutide, a DPP IV-resistant GLP-1 dimer bound to albumin was approved by both FDA and European authorities in 2014. FDA-approved dulaglutide is a GLP-1 dimer linked to human immunoglobulin G4. Their half-lifes, 4 – 7 days, are in the same range as the half-life of the recently developed GLP-1 analog semaglutide. These peptides are considerably more stable than exenatide.
- DPP IV inhibitors represent another approach for the treatment of diabetes. Sitagliptin is the first candidate of this novel class of antihyperglycemic agents that has been approved by the FDA. Alogliptin, linagliptin, saxagliptin, and vildagliptin have been approved in the USA and in various countries worldwide. These DPP IV inhibitors can be used either alone or in combination with other oral antihyperglycemic agents (such as metformin or a thiazolidine-dione) for treatment of diabetes mellitus type 2.
- Pramlintide, a soluble amylin analog, has gained FDA approval as an adjunct to insulin therapy in type 1 and type 2 diabetes. Like amylin, pramlintide acts centrally and decreases glucagon secretion, slows gastric emptying and induces satiety.
- Insulin therapy is also indicated in the treatment of type 2 diabetes for the management of severe hyperglycemia after failure of oral agents. Long-acting insulin analogs could be obtained by replacement of Asn21 (A-chain) by glycine and addition of two arginine residues (insulin glargine) or acylation of lysine29 (B-chain) with long-chain fatty acids. Insulin degludec is obtained by palmitoylation, which distinctly improves the metabolic stability of the peptide hormone by hexamer formation. Insulin degludec was approved in Europe in 2015 for treatment of children and adolescents with diabetes.
- C-Peptide is biologically active. Clinical studies showed that administration of C-peptide to diabetes type 1 patients lackingthe peptide alleviates nerve and renal dysfunctions associated with the disease. A long-acting analog, PEGylated C-peptide, is under evaluation.
According to the WHO, the number of people living with diabetes has almost quadrupled since 1980 to 422 million adults.
Prospects
Although some of the agents described above are still in the early phases of investigation, there is little doubt that the therapy of diabetes will undergo major changes in the near future. It is important to diagnose all type 2 diabetics at an earlier stage (for example by making self monitoring of blood glucose easier) and begin treatment in an attempt to minimize the diabetes-associated complications.
The identification of the genetic components of type 1 and type 2 diabetes is an important area of research, because elucidation of the diabetes genes will influence all efforts towards an understanding of the disease, its complications, and its treatment, cure, and prevention. For this reason, genomic DNA from subjects with severe insulin resistance has been screened for mutations in genes that are implicated in insulin signaling. Indeed, a mutation in the gene encoding the serine/threonine kinase AKT2 (also known as PKBβ) could be identified.
In the 1970s, the insulin receptor was discovered, and 10 years later, its tyrosine kinase activity was demonstrated.
AKT2 is highly expressed in insulin-sensitive tissues and has been implicated in insulin-regulated glucose uptake into muscle and fat cells by promoting the translocation of glucose transporter 4 (GLUT4) to the cell surface. Advances in genomics, proteomics and metabolomics will help us to further understandthe causes of type 1 and type 2 diabetes and could eventually lead to novel therapeutic approaches.
Subscribe to our newsletter
"*" indicates required fields
References
R.J. Mahler and M.L. Adler
Clinical Review 102: Type 2 diabetes mellitus: update on diagnosis, pathophysiology, and treatment.
J. Clin. Endocrinol. Metab. 84, 1165 -1171 (1999)
L.B. Knudson et al.
GLP-1 derivatives as novel compounds for the treatment of type 2 diabetes: selection of NN2211 for clinical development.
Drugs Future 26, 677-685 (2001)
A.D. Baron et al.
Novel peptides under development for the treatment of type 1 and type 2 diabetes mellitus. Curr. Drug Targets Immune Endocr.
Metabol. Disord. 2, 63-82 (2002)
D.J. Drucker
Biological actions and therapeutic potential of the glucagon-like peptides.
Gastroenterology 122, 531-544 (2002)
J.J. Holst
Gastric inhibitory polypeptide analogues: do they have a therapeutic role in diabetes mellitus similar to that of glucagon-like peptide-1?
BioDrugs 16, 175-181 (2002)
J.J. Holst
Therapy of type 2 diabetes mellitus based on the actions of glucagonlike peptide-1.
Diabetes Metab. Res. Rev. 18, 430-441 (2002)
S. Yakar et al.
The role of circulating IGF-I. Lessons from human and animal models.
Endocrine 19, 239-248 (2002)
B. Ahren
Gut peptides and type 2 diabetes mellitus treatment.
Current Diabetes Reports 3, 365-372 (2003)
D.J. Drucker
Enhancing incretin action for the treatment of type 2 diabetes.
Diabetes Care 26, 2929-2940 (2003)
J.M. Egan et al.
GLP-1 receptor agonists are growth and differentiation factors for pancreatic islet beta cells.
Diabetes Metab. Res. Rev. 19, 115-123 (2003)
R.I.G. Holt et al.
The role of the growth hormoneinsulin-like growth factor axis in glucose homeostasis.
Diabet. Med. 20, 3-15 (2003)
G.G. Holz and O.G. Chepurny
Glucagon-like peptide-1 synthetic analogs: new therapeutic agents for use in the treatment of diabetes mellitus.
Curr. Med. Chem. 10, 2471-2483 (2003)
G. Jiang and B.B. Zhang
Glucagon and regulation of glucose metabolism.
Am. J. Physiol. Endocrinol. Metab. 284, E671-E678 (2003)
T.J. Kieffer
GIP or not GIP? That is the question.
Trends Pharmacol. Sci. 24, 110-112 (2003)
L. Marzban et al.
Islet amyloid polypeptide and type 2 diabetes.
Exp. Gerontol. 38, 347-351 (2003)
P.G. McTernan et al.
Resistin and type 2 diabetes: regulation of resistin expression by insulin and rosiglitazone and the effects of recombinant resistin on lipid and glucose metabolism in human differentiated adipocytes.
J. Clin. Endocrinol. Metab. 88, 6098- 6106 (2003)
J.J. Meier et al.
Glucagon-like peptide 1 and gastric inhibitory polypeptide: potential applications in type 2 diabetes mellitus.
BioDrugs 17, 93-102 (2003)
N. Moller et al.
Effects of ageing on insulin secretion and action.
Horm. Res. 60, 102-104 (2003)
T. Perry and N.H. Greig
The glucagon-like peptides: a double-edged therapeutic sword?
Trends Pharmacol. Sci. 24, 377-383 (2003)
B. Thorens
Gluco-incretin hormones in insulin secretion and diabetes.
Med. Sci. (Paris) 19, 860-863 (2003)
T. Vahl and D. D’Alessio
Enteroinsular signaling: perspectives on the role of the gastrointestinal hormones glucagon-like peptide 1 and glucose-dependent insulinotropic polypeptide in normal and abnormal glucose metabolism.
Curr. Opin. Clin. Nutr. Metab. Care 6, 461-468 (2003)
M.F. White
Insulin signaling in health and disease.
Science 302, 1710-1711 (2003)
P.E. Wiedeman and J.M. Trevillyan
Dipeptidyl peptidase IV inhibitors for the treatment of impaired glucose tolerance and type 2 diabetes.
Curr. Opin. Investig. Drugs 4, 412-420 (2003)
Y. Dor et al.
Adult pancreatic β-cells are formed by self-duplication rather than stem-cell differentiation.
Nature 429, 41-46 (2004)
E.A. Gale et al.
European Nicotinamide Diabetes Intervention Trial (ENDIT) Group: A randomised controlled trial of intervention before the onset of type 1 diabetes.
Lancet 363, 925-931 (2004)
S. George et al.
A family with severe insulin resistance and diabetes due to a mutation in AKT2.
Science 304, 1325-1328 (2004)
J.F. List and J.F. Habener
Glucagon-like peptide 1 agonists and the development and growth of pancreatic beta-cells.
Am. J. Physiol. Endocrinol. Metab. 286, E875-E881 (2004)
L.L. Nielsen et al.
Pharmacology of exenatide (synthetic exendin-4): a potential therapeutic for improved glycemic control of type 2 diabetes.
Regul. Peptides 117, 77-88 (2004)
A. Nourparvar et al.
Novel strategies for the pharmacological management of type 2 diabetes.
Trends Pharmacol. Sci. 25, 86-91 (2004)
S.M. Rangwala and M.A. Lazar
Peroxisome proliferator-activated receptor γ in diabetes and metabolism.
Trends Pharmacol. Sci. 25, 331-336 (2004)
Y. Sato et al.
C-peptide fragments stimulate glucose utilization in diabetic rats.
Cell. Mol. Life Sci. 61, 727-732 (2004)
T.P. Vahl and D.A. D‘Alessio
Gut peptides in the treatment of diabetes mellitus.
Expert. Opin. Investig. Drugs. 13, 177- 188 (2004)
J.F. Gautier et al.
Biological actions of the incretins GIP and GLP-1 and therapeutic perspectives in patients with type 2 diabetes.
Diabetes Metab. 31, 233-242 (2005)
E.M. Sinclair and D.J. Drucker
Proglucagon-derived peptides: mechanisms of action and therapeutic potential.
Physiology 20, 357-365 (2005)
D.J. Drucker and M.A. Nauck
The incretin system: glucagon-like peptide-1 receptor agonists and dipeptidyl peptidase-4 inhibitors in type 2 diabetes.
Lancet 368, 1696-1705 (2006)
K. Fujioka
Pathophysiology of type 2 diabetes and the role of incretin hormones and beta-cell dysfunction.
JAAPA Suppl, 3-8 (2007)
R. Gadsby
New treatments for type 2 diabetes – the DPP4 inhibitors.
Prim. Care Diabetes 1, 209-211 (2007)
J. Meece
Pancreatic islet dysfunction in type 2 diabetes: a rational target for incretin-based therapies.
Curr. Med. Res. Opin. 23, 933-944 (2007)
J. Wahren et al.
C-Peptide is a bioactive peptide.
Diabetologia 50, 503-509 (2007)
A.M. Abbatecola et al.
New approaches to treating type 2 diabetes mellitus in the elderly: role of incretin therapies.
Drugs Aging 25, 913-925 (2008)
S.E. Inzucchi and D.K. McGuire
New drugs for the treatment of diabetes: part II: Incretin-based therapy and beyond.
Circulation 117, 574-584 (2008)
A.M. Lambeir et al.
DPP4 inhibitors for diabetes – what next?
Biochem. Pharmacol. 76, 1637-1643 (2008)
N. Mikhail
Incretin mimetics and dipeptidyl peptidase 4 inhibitors in clinical trials for the treatment of type 2 diabetes.
Expert Opin. Investig. Drugs 17, 845-853 (2008)
Y. Ng et al.
Rapid activation of Akt2 is sufficient to stimulate GLUT4 translocation in 3T3-L1 adipocytes.
Cell. Metab. 7, 348-356 (2008)
P.R. Flatt et al.
Recent advances in antidiabetic drug therapies targeting the enteroinsular axis.
Curr. Drug Metab. 10, 125-137 (2009)
R. Gupta et al.
Emerging drug candidates of dipeptidyl peptidase IV (DPP IV) inhibitor class for the treatment of type 2 diabetes.
Curr. Drug Targets 10, 71-87 (2009)
F.K. Knop et al.
Incretin-based therapy of type 2 diabetes mellitus.
Curr. Protein Pept. Sci. 10, 46-55 (2009)
T. Salvatore et al.
Progress in the oral treatment of type 2 diabetes: update on DPP-IV inhibitors.
Curr. Diabetes Rev. 5, 92-101 (2009)
W. Benalla et al.
Antidiabetic medicinal plants as a source of alpha glucosidase inhibitors.
Curr. Diab. Rev. 6 247-254 (2010)
M. Christensen and F.K. Knop
Once-weekly GLP-1 agonists: How do they differ from exenatide and liraglutide?
Curr. Diab. Rep. 10, 124-132 (2010)
B. Gallwitz
Benefit-risk assessment of exenatide in the therapy of type 2 diabetes mellitus.
Drug Saf. 33, 87-100 (2010)
J. Gerich
DPP-4 inhibitors: what may be the clinical differentiators?
Diabetes Res. Clin. Pract. 90, 131-140 (2010)
Y. Seino et al.
GIP and GLP-1, the two incretin hormones: Similarities and differences.
J. Diabetes Investig. 1, 8-23 (2010)
B. Ahren
The future of incretin-based therapy: novel avenues – novel targets.
Diabetes Obes. Metab. 13 Suppl 1, 158-166 (2011)
M.J. Davies et al.
Liraglutide – overview of the preclinical and clinical data and its role in the treatment of type 2 diabetes.
Diabetes Obes. Metab. 13, 207-220 (2011)
B. Gallwitz
GLP-1 agonists and dipeptidyl-peptidase IV inhibitors.
Handb. Exp. Pharmacol. 53-74 (2011)
D.M. Irwin and K.J. Prentice
Incretin hormones and the expanding families of glucagon-like sequences and their receptors.
Diabetes Obes. Metab. 13 Suppl 1, 69-81 (2011)
G.J. Ryan and Y. Hardy
Liraglutide: once-daily GLP-1 agonist for the treatment of type 2 diabetes.
J. Clin. Pharm. Ther. 36, 260-274 (2011)
J. Shubrook et al.
Saxagliptin: A Selective DPP-4 Inhibitor for the Treatment of Type 2 Diabetes Mellitus.
Clin. Med. Insights Endocrinol. Diabetes 4, 1-12 (2011)
P. Westermark et al.
Islet amyloid polypeptide, islet amyloid, and diabetes mellitus.
Physiol. Rev. 91, 795-826 (2011)
K. Hunter and C. Holscher
Drugs developed to treat diabetes, liraglutide and lixisenatide, cross the blood brain barrier and enhance neurogenesis.
BMC Neurosci. 13, 33 (2012)
M.A. Jackson et al.
Stable liquid glucagon formulations for rescue treatment and bi-hormonal closed-loop pancreas.
Curr. Diab. Rep. 12, 705-710 (2012)
N.R. Pinelli et al.
Exogenous glucagon-like peptide-1 for hyperglycemia in critically ill patients.
Ann. Pharmacother. 46, 124-129 (2012)
A. Pocai
Unraveling oxyntomodulin, GLP1’s enigmatic brother.
J. Endocrinol. 215, 335-346 (2012)
B.M. Wice et al.
Xenin-25 amplifies GIP-mediated insulin secretion in humans with normal and impaired glucose tolerance but not type 2 diabetes.
Diabetes 61, 1793-1800 (2012)
S. Calanna et al.
Secretion of Glucose-Dependent Insulinotropic Polypeptide in Patients With Type 2 Diabetes.
Diabetes Care 36. 3346-3352 (2013)
G. Grunberger
Novel therapies for the management of type 2 diabetes mellitus: part 1. pramlintide and bromocriptine-QR.
J. Diabetes 5,110-117 (2013)
International Diabetes Federation
IDF Diabetes Atlas (6th ed.)
(2013)
N. Irwin and P.R. Flatt
Enteroendocrine hormone mimetics for the treatment of obesity and diabetes.
Curr. Opin. Pharmacol. 13, 989-995 (2013)
K. Pillai and P. Govender
Amylin uncovered: a review on the polypeptide responsible for type II diabetes.
Biomed. Res. Int. 2013, 826706 (2013)
D. Bataille and S. Dalle
The forgotten members of the glucagon family.
Diabetes Res. Clin. Pract. 106,1-10 (2014)
S.L. Booth et al.
Bone as an endocrine organ relevant to diabetes.
Curr. Diab. Rep. 14, 556 (2014)
C. Chakraborty et al.
Understanding the molecular dynamics of type-2 diabetes drug target DPP-4 and its interaction with Sitagliptin and inhibitor Diprotin-A.
Cell. Biochem. Biophys. 70, 907-922 (2014)
S. Chowdhury et al.
Xenin-25 delays gastric emptying and reduces postprandial glucose levels in humans with and without type 2 diabetes.
Am. J. Physiol. Gastrointest. Liver Physiol. 306, G301-G309 (2014)
R.E. George and S. Joseph
A review of newer treatment approaches for type-2 diabetes: Focusing safety and efficacy of incretin based therapy.
Saudi Pharm. J. 22, 403-410 (2014)
C.F. Godfredsen et al.
The human GLP-1 analogs liraglutide and semaglutide: absence of histopathological effects on the pancreas in nonhuman primates.
Diabetes 6. 2486-2497 (2014)
A.F. Godoy-Matos
The role of glucagon on type 2 diabetes at a glance.
Diabetol. Metab. Syndr. 6, 91 (2014)
L. Juillerat-Jeanneret
Dipeptidyl peptidase IV and its inhibitors: therapeutics for type 2 diabetes and what else?
J. Med. Chem. 57, 2197-2212 (2014)
S.E. Kahn et al.
Pathophysiology and treatment of type 2 diabetes: perspectives on the past, present, and future.
Lancet 383, 1068-1083 (2014)
R.W. Nelson and C.E. Reusch
Animal models of disease: classification and etiology of diabetes in dogs and cats.
J. Endocrinol. 222, T1-T9 (2014)
A. Pocai
Action and therapeutic potential of oxyntomodulin.
Mol. Metab. 3, 241-251 (2014)
M. Rigato and G.P. Fadini
Comparative effectiveness of liraglutide in the treatment of type 2 diabetes.
Diabetes Metab. Syndr. Obes. 7, 107- 120 (2014)
G. Umpierrez et al.
Efficacy and safety of dulaglutide monotherapy versus metformin in type 2 diabetes in a randomized controlled trial (AWARD-3).
Diabetes Care 37, 2168-2176 (2014)
H.A. Blair and G.M. Keating
Albiglutide: a review of its use in patients with type 2 diabetes mellitus.
Drugs 75, 651-663 (2015)
D.L. Hay et al.
Amylin: Pharmacology, Physiology, and Clinical Potential.
Pharmacol. Rev. 67, 564-600 (2015)
I. Kanazawa
Osteocalcin as a hormone regulatingglucose metabolism.
World J. Diabetes 6,1345-1354 (2015)
J. Lau et al.
Discovery of the Once-Weekly Glucagon-Like Peptide-1 (GLP-1)
Analogue Semaglutide. J. Med. Chem. 58, 7370-7380 (2015)
C. Liu et al.
N-Acetyl Cysteine improves the diabetic cardiac function: possible role of fibrosis inhibition.
BMC Cardiovasc. Disord. 15, 84 (2015)
F. Mittermeyer et al.
Addressing unmet medical needs in type 2 diabetes: a narrative review of drugs under development.
Curr. Diabetes Rev. 11, 17-31 (2015)
D.A. Sandoval and D.A. D’Alessio
Physiology of proglucagon peptides: role of glucagon and GLP-1 in health and disease.
Physiol. Rev. 95, 513-548 (2015)
J.A. Shaw et al.
C-peptide as a Therapy for Kidney Disease: A Systematic Review and Meta-Analysis.
PLoS ONE 10, e0127439 (2015)
M. Visa et al.
Islet amyloid polypeptide exerts a novel autocrine action in beta-cell signaling and proliferation.
FASEB J. 29, 2970-2979 (2015)
L. Whiting et al.
Glicentin-related pancreatic polypeptide inhibits glucose-stimulated insulin secretion from the isolated pancreas of adult male rats.
Physiol. Rep. 3 (2015)
L. Zhong et al.
Recent Advances in Dipeptidyl-Peptidase-4 Inhibition Therapy: Lessons from the Bench and Clinical Trials.
J. Diabetes Res. 606031 (2015)
S.L. Anderson and J.M. Trujillo
Lixisenatide in type 2 diabetes: latest evidence and clinical usefulness.
Ther. Adv. Chronic Dis. 7, 4-17 (2016)
O. Bouttefeux et al.
Delivery of Peptides Via the Oral Route: Diabetes Treatment by Peptide-Loaded Nanoparticles.
Curr. Pharm. Des. 22, 1161-1176 (2016)
S. Kalra et al.
Glucagon-like peptide-1 receptor agonists in the treatment of type 2 diabetes: Past, present, and future.
Indian J. Endocrinol. Metab. 20, 254-267 (2016)
C.Y. Lee
Glucagon-Like Peptide-1 Formulation – the Present and Future Developmentin Diabetes Treatment.
Basic Clin. Pharmacol. Toxicol. 118, 173-180 (2016)
L. Li et al.
Dipeptidyl peptidase-4 inhibitors and risk of heart failure in type 2 diabetes: systematic review and meta-analysis of randomised and observational studies.
BMJ 352, i610 (2016)
M.G. Minze and L.M. Chastain
Combination therapies in the management of type 2 diabetes: the use of insulin degludec/liraglutide.
Ther. Clin. Risk Manag. 12, 471-478 (2016)
R.G. Mirmira et al.
Biomarkers of beta-Cell Stress and Death in Type 1 Diabetes.
Curr. Diab. Rep. 16, 95 (2016)
A.K. Sharma et al.
Albiglutide: Is a better hope against diabetes mellitus?
Biomed. Pharmacother. 77, 120-128 (2016)
L. Sobrevia et al.
Insulin Is a Key Modulator of Fetoplacental Endothelium Metabolic Disturbances in Gestational Diabetes Mellitus.
Front. Physiol. 7, 119 (2016)
World Health Organization
Diabetes (2022), World Health Organization, Retrieved 2022, from https://www.who.int/health-topics/diabetes#tab=tab_1